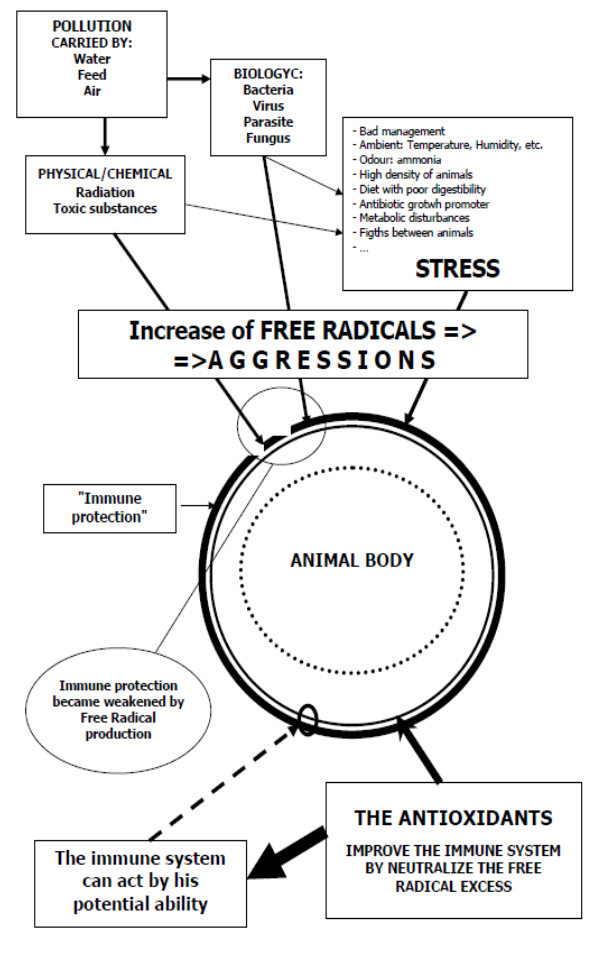
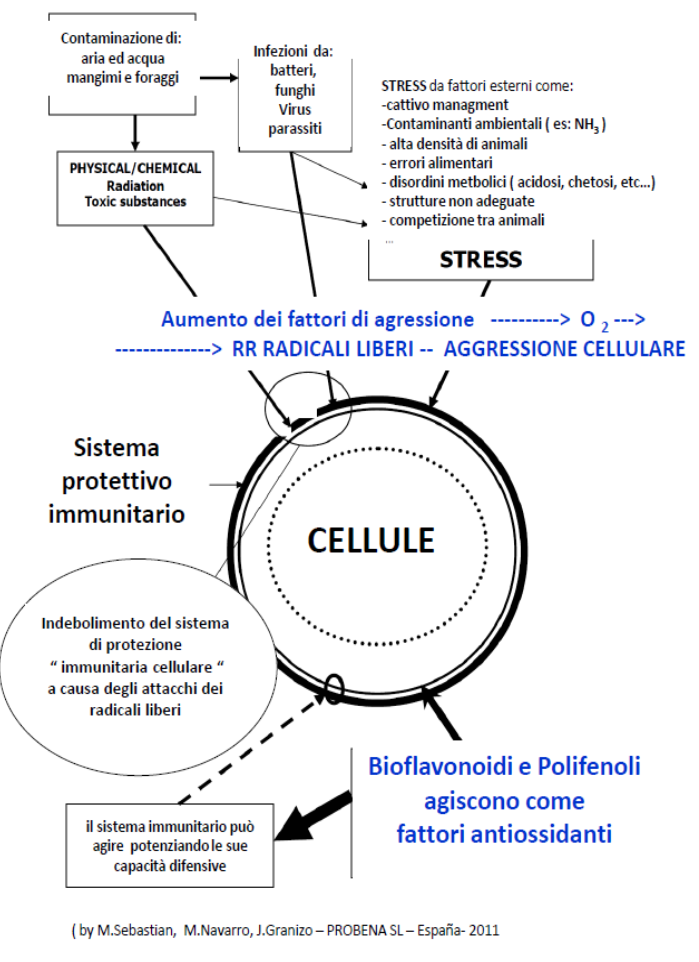
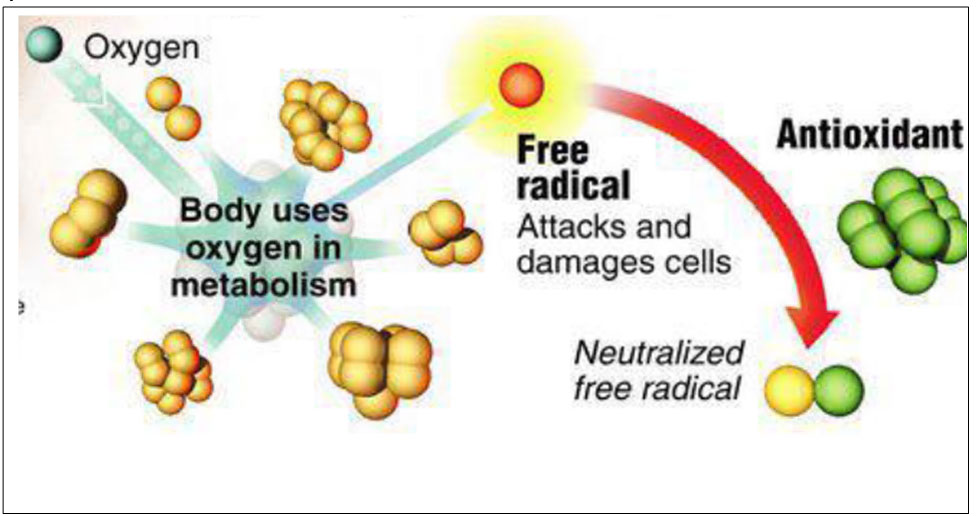
Il ruolo e l’utilizzo degli antiossidanti in nutrizione e salute animale – Terza parte
Sistemi di valutazione della capacità antiossidante di un alimento
I sistemi per valutare la capacità potenziale di agire come antiossidante di un dato alimento, ovvero di interferire sul sistema metabolico e immunitario degli animali, sono praticamente due:
- La capacità antiossidante equivalente TEAC ( in Trolox ) ovvero la capacità antiossidante equivalente di Trolox (TEAC) che misura la capacità antiossidante di una particolare sostanza, rispetto allo standard, Trolox o (6-idrossi-2,5,7,8-tetrametilchroman-2-acido carbossilico). È un analogo solubile in acqua di Vitamina E. La capacità antiossidante equivalente di Trolox (TEAC) è una misurazione della resistenza antiossidante basata su Trolox, misurata in unità chiamata Trolox micromol ITE, ad es. micromol / 100 g ( esame in vitro ). A causa delle difficoltà di misurare le singole componenti antiossidanti di una miscela complessa (es: come gli agrumi, i mirtilli , pomodori, tè verde, etc…), l’equivalenza di Trolox è, oggi, utilizzata come riferimento per la capacità antiossidante di tale miscela. L’equivalenza di Trolox è spesso misurata utilizzando la capacità antiossidante di alimenti ( alimenti ricchi di polifenoli inclusi quelli ad uso zootecnico ) come nella capacità di riduzione del ferro del plasma (FRAP). Tale test è eseguito in vitro e misura esclusivamente la capacità antiossidante potenziale dell’alimento come valore standard, senza verificarne la sua attività a livello metabolico. Questo metodo esprime una scala di valori detta ORAC ( Oxigen Radicals Absorbance Capacity )
- Test KRL ( by M. Prost , bretto Kirial – Spirial – Brevetto M. PROST/SPIRAL – Octobre 2003)) Test KRL invece fornisce invece una misura dello stato delle difese anti-radicali globali di un soggetto e determina le potenzialità difensive nei confronti dei radicali liberi di vari prodotti ( vitamine, alimenti, spezie, etc…). Il test in pratica simula un attacco ossidativo “ tip” ai globuli rossi in un ambiente controllato e standardizzato.
Esempio di scala ORAC
Quando si confrontano i dati ORAC ( TERAC )occorre prestare attenzione affinché le unità e il cibo che si confronta siano simili. Alcune valutazioni, infatti, dovranno essere valutate per unità ORAC per grammo di peso secco del cibo intero fresco ( es: frutta ) o del frutto secco macinato o congelato. In ogni valutazione, i cibi diversi possono apparire con valori più alti di ORAC, è necessario pertanto rapportarli con gli stessi parametri ( secco, disidratato e/o intero, etc….) ( es: allo stesso modo, il grande contenuto di acqua nell’anguria può far apparire come se questo frutto fosse basso in ORAC. cosa invece non esatta).Allo stesso modo, occorre prendere in considerazione la tipica quantità di alimento utilizzato per erbe e spezie applicando la scala ORAC, ma in quantità molto più basse dal momento che si parla di concentrati di alimenti interi intatti.
Oggigiorno numerose aziende e marketing di prodotti alimentari e di bevande dietetiche e premix per animali, etc… hanno erroneamente capitalizzato il loro rating ORAC promuovendo i prodotti dichiarati “alti in ORAC o TEAC”. Poiché la maggior parte di questi valori ORAC non sono stati convalidati in modo oggettivo da Enti e/o Istituzioni indipendenti o sottoposti a revisioni parziali per la pubblicazione in letteratura scientifica, in molti casi rimangono non confermati e non scientificamente credibili e possono indurre in errore gli utilizzatori. Il Dipartimento dell’Agricoltura degli Stati Uniti (USA) ha ritirato la sua affidabilità nel 2012 come biologicamente non validi, affermando che “ i dati relativi alla capacità antiossidante degli alimenti generati da in vitro (test- tube) non possono essere estrapolati agli effetti in vivo (sia per persone che per animali) e le sperimentazioni cliniche per testare i benefici degli antiossidanti dietetici hanno prodotto non affidabili”.
Test KRL di resistenza ai fattori ossidanti (by Dr. Michel Prost – SPIRAL)
Test KRL fornisce una misura dello stato delle difese anti-radicali globali di un soggetto e determina le potenzialità difensive nei confronti dei radicali liberi di vari prodotti ( vitamine, antiossidanti, alimenti o mangimi, etc…)
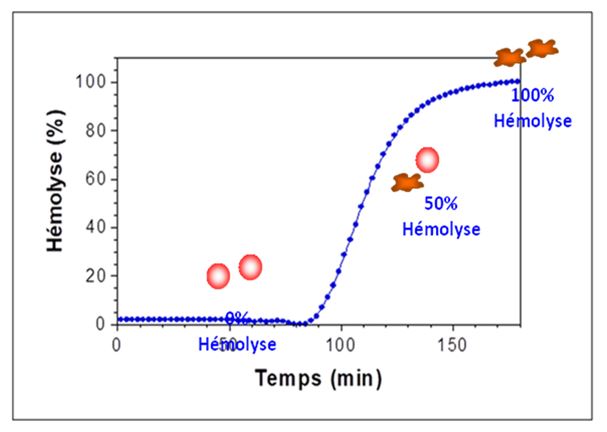
Il test in pratica simula un attacco ossidativo “ tipo” ai globuli rossi in un ambiente controllato e standardizzato. In queste condizioni, gli eritrociti non vengono influenzati da altri fattori enzimatici e strutture molecolare per sopportare l’attacco ossidativo fino a quando la membrana cellulare si altera al punto tale da perdere il loro contenuto cellulare. La resistenza degli eritrociti sottoposti a test è pertanto espressa dal tempo impiegato per rilasciare il 50% del contenuto di emoglobina.
L’importanza medico-scientifica di questo test è che esso riflette la capacità globale di difesa di un individuo nei confronti dell’aggressione dei radicali liberi coinvolti in molte malattie e che ci permette di valutare preventivamente la capacità “ potenziale” di difesa dell’individuo in un habitat che tenga conto di tutti i fattori positivi come vitamine, capacità della difesa enzimatica …) e fattori ( stress, cattiva alimentazione, micotossine, ecc).
METODO PER EFFETTUARE IL TEST KRL ( brevetto Prost- Spiral )
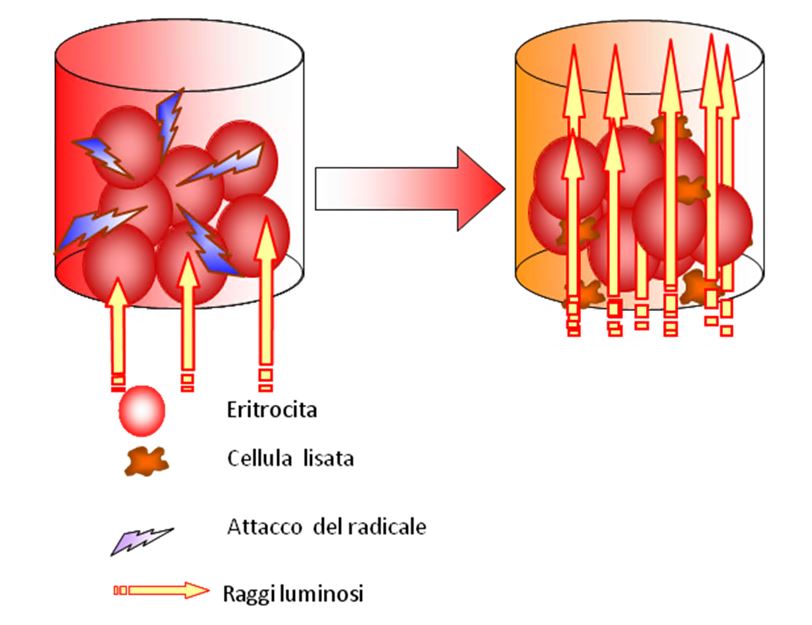
Applicazione del test KRL
È stata valutata l’applicazione del kit Radicaux Libres (KRL) per valutare l’attività antiossidante del sangue totale nei suini. Il KRL è stato convalidato anche dall’FDA – USA e EFSA – Europeo ed è oggi considerato il test più affidabile ed ampiamente utilizzato negli esseri umani per valutare l’efficacia dei trattamenti naturali o farmaceutici per valutare le attività antiossidanti naturali( polifenoli e bioflavonoidi) o sintetici ( vitamina E e Vitamina C). Il test viene raccomandato come uno strumento efficace per valutare l’attività antiossidante degli ingredienti alimentari negli alimenti per suini ( by Rossi R, Pastorelli G, Corino C- Res Vet Sci 2013 Apr ).
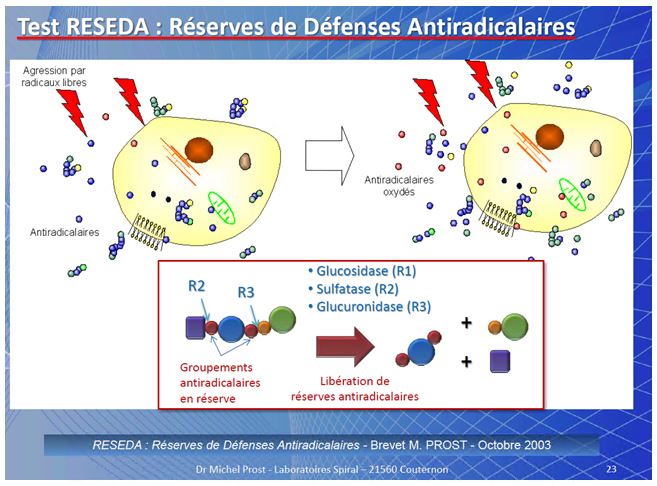
Il principio del RESEDA (Réserves de Défenses Antiradicalaires – Brevetto M. PROST/SPIRAL – Octobre 2003)
È stato dimostrato che le cellule sottoposte allo stress metabolico dei radicali liberi hanno la capacità di aumentare i loro sistemi di difesa cellulare accumulando un potenziale di difesa contro gli stessi che utilizzano in caso di necessita (stress ossidativo ). Questo potenziale cambia a seconda delle condizioni fisiologiche in cui si trova l’organismo e a seconda della quantità di anti radicali liberi (in pratica antiossidanti) che la cellula è riuscita ad accumulare. Tale principio (RESEDA) in pratica, dimostra la capacità di “auto-difesa cellulare” utilizzando gli antiossidanti accumulati (principio brevettato dal Dr. Michel Prost/ Spiral Ottobre 2003).
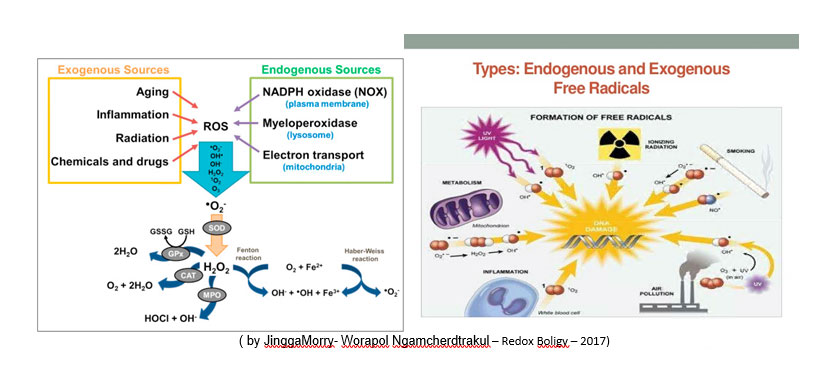
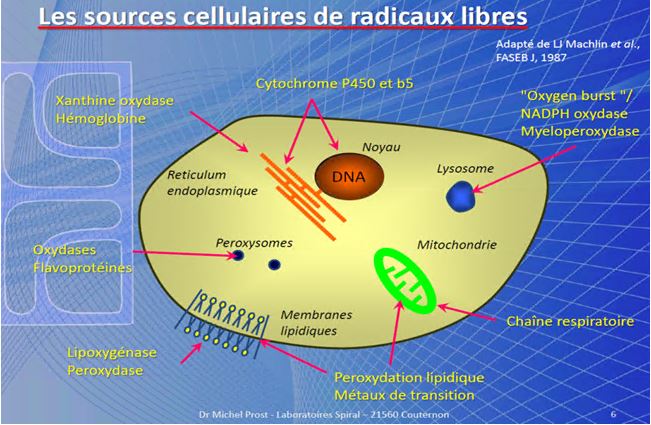
FONTI DI RADICALI LIBERI( by M.Prost )
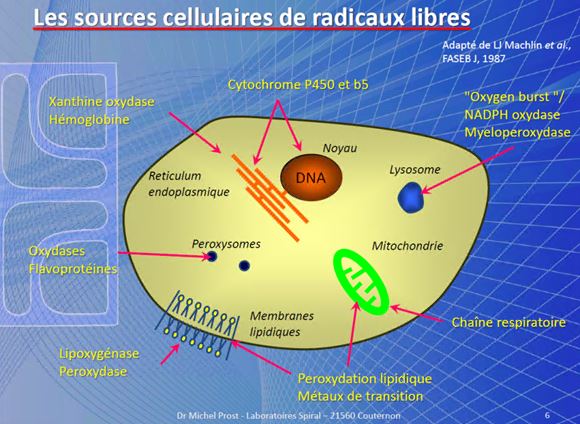
LE DIFESE CELLULARI CONTRO I RADICALI LIBERI( by M.prost )
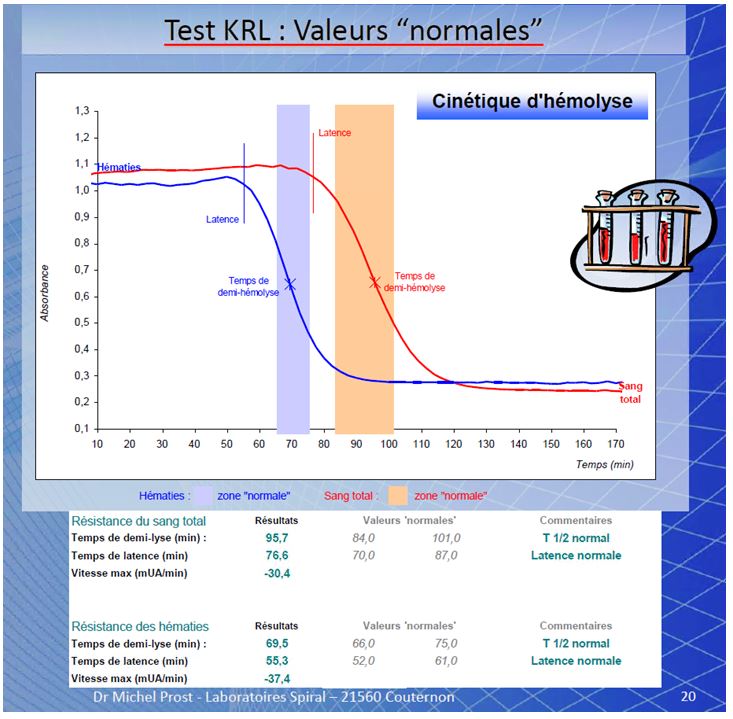
CINETICA DELL’ EMOLISI ( by M.prost )
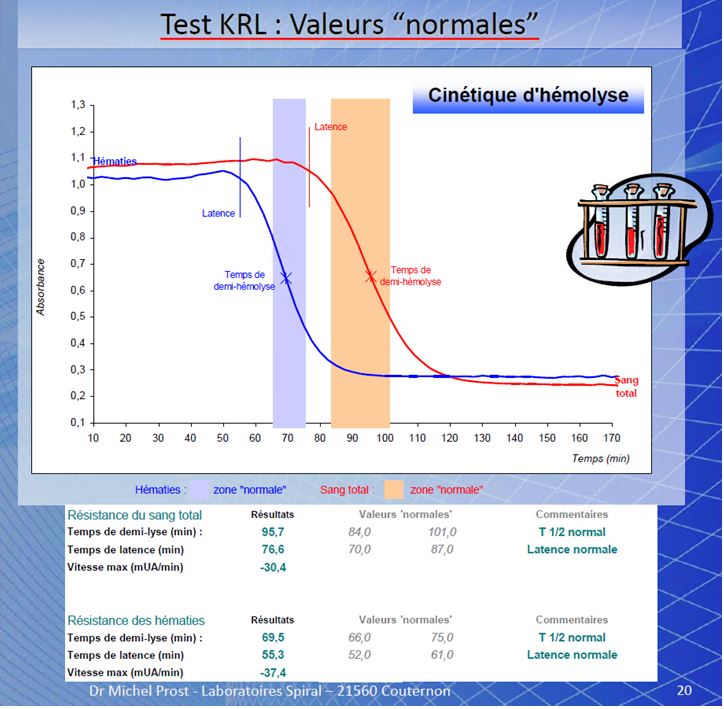
Risultati su sangue di soggetti trattati con antiossidanti
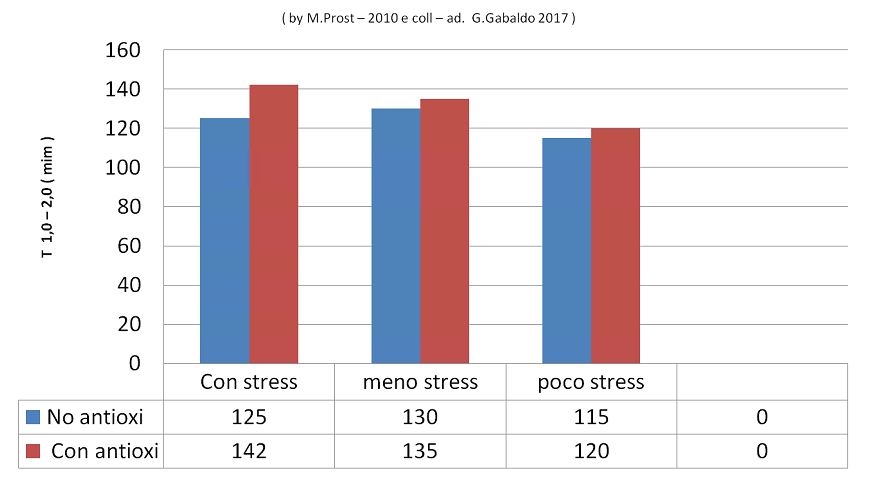
TEST ESEGUITO SU DEI SUINI TRATTARI E NON TRATTATI CON ANTIOSSIDANTI ( by M.prost )
Riconoscimento Giuridico a livello Europeo del test KRL